Samo upakowanie DNA w jądrze komórkowym nie jest jedynie problemem miejsca. Komórka musi zmieścić około 2 metrów DNA w przestrzeni liczonej w mikrometrach, a jednocześnie w każdej chwili wiedzieć, które fragmenty mają być odczytane, skopiowane albo naprawione. Dlatego ten temat najlepiej rozumieć jako połączenie biologii strukturalnej, regulacji genów i organizacji całego jądra.
Najważniejsze fakty o organizacji DNA w jądrze
- Podstawową jednostką upakowania jest nukleosom, czyli DNA owinięte wokół oktameru histonów.
- Luźniej upakowana chromatyna zwykle sprzyja aktywności genów, a ciaśniej upakowana ogranicza dostęp do DNA.
- Na wyższym poziomie liczą się pętle chromatynowe, domeny kontaktów i powiązanie z otoczką jądrową.
- Organizacja DNA jest dynamiczna: zmienia się podczas transkrypcji, replikacji i naprawy uszkodzeń.
- Błędy w tej architekturze mogą prowadzić do zaburzeń rozwoju, niestabilności genomu i nowotworów.
Jak DNA mieści się w jądrze komórkowym
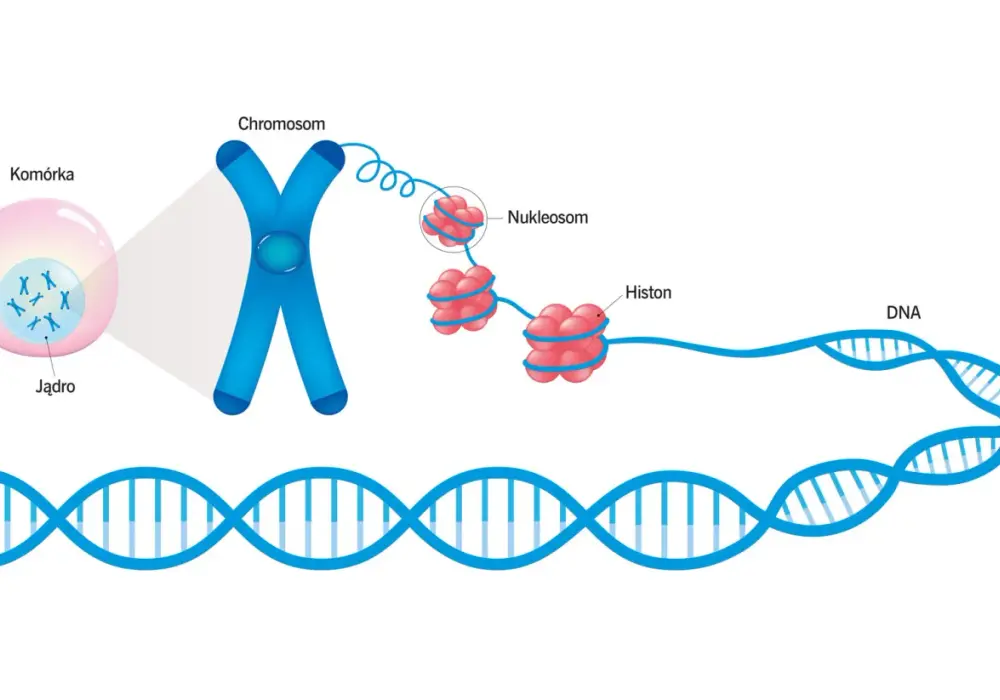
Gdy tłumaczę ten temat, zaczynam od prostej skali. Ludzka komórka somatyczna zawiera bardzo długi łańcuch DNA, a jądro jest od niego nieporównanie mniejsze. Komórka radzi sobie z tym nie przez przypadkowe „zwijanie”, ale przez bardzo uporządkowaną sekwencję poziomów organizacji.
Na pierwszym poziomie stoi nukleosom. To odcinek DNA owinięty wokół oktameru histonów, czyli dwóch kopii białek H2A, H2B, H3 i H4. Wokół tego rdzenia nawinięte jest około 147 par zasad, a kolejne fragmenty DNA łącznikowego zwiększają długość całej jednostki. Histon H1 pomaga stabilizować ten układ i porządkuje dalsze zagęszczanie materiału genetycznego.
| Poziom organizacji | Przybliżona skala | Co się dzieje |
|---|---|---|
| Podwójna helisa DNA | ok. 2 nm | Podstawowa cząsteczka nośna informacji genetycznej |
| Nukleosom | ok. 10 nm | DNA owija się wokół histonów i zaczyna tworzyć chromatynę |
| Pętle i domeny chromatynowe | zmienna | Fragmenty DNA organizują się przestrzennie w kontakty lokalne i dalekie |
| Chromosom metafazowy | najsilniejsza kondensacja | Materiał genetyczny jest maksymalnie upakowany przed podziałem komórki |
W starszych schematach często pokazuje się jeszcze sztywne włókno 30 nm jako kolejny obowiązkowy etap. Ja podchodzę do tego ostrożnie, bo w żywych komórkach nie jest to uniwersalny, stały model dla wszystkich typów tkanek i wszystkich warunków. Lepiej myśleć o chromatynie jako o układzie dynamicznych zagęszczeń, który zależy od stanu komórki, a nie o jednej, z góry ustalonej rurce upakowania. I właśnie ta elastyczność prowadzi do pytania, dlaczego część DNA jest łatwo dostępna, a część pozostaje schowana.
Dlaczego dostępność chromatyny zmienia pracę genów
Sam stopień upakowania nie mówi jeszcze wszystkiego. Dla biologii kluczowe jest to, jak łatwo białka regulacyjne mogą dotrzeć do DNA. W praktyce oznacza to różnicę między chromatyną bardziej otwartą a bardziej zwartą, czyli między euchromatyną i heterochromatyną.
| Cecha | Euchromatyna | Heterochromatyna |
|---|---|---|
| Upakowanie | Luźniejsze | Ciaśniejsze |
| Dostęp do DNA | Łatwiejszy | Trudniejszy |
| Typowy efekt | Aktywna transkrypcja i szybka odpowiedź komórki | Wyciszenie lub stabilizacja trudniej dostępnych regionów |
| Znaczenie biologiczne | Umożliwia odczyt genów potrzebnych tu i teraz | Pomaga utrzymać porządek genomu i kontrolować nadmiar aktywności |
To rozróżnienie bywa upraszczane, ale nie powinno być rozumiane mechanicznie. Heterochromatyna nie oznacza „martwego DNA”, a euchromatyna nie jest chaotycznie rozlazła. Wiele zależy od modyfikacji histonów i białek, które te modyfikacje odczytują. Acetylacja histonów zwykle sprzyja rozluźnieniu chromatyny, natomiast metylacja może działać różnie w zależności od miejsca i kontekstu. Z punktu widzenia komórki ważny jest nie tylko sam poziom upakowania, ale też dostępność i czasowy rytm otwierania fragmentów DNA.
W praktyce właśnie tak działa regulacja genów: komórka nie wyłącza wszystkiego naraz, tylko precyzyjnie odsłania te obszary, które są potrzebne w danym momencie. Na tym tle łatwo przejść do jeszcze wyższego poziomu organizacji, czyli do przestrzennej architektury całego jądra.
Jak komórka porządkuje DNA w trzech wymiarach
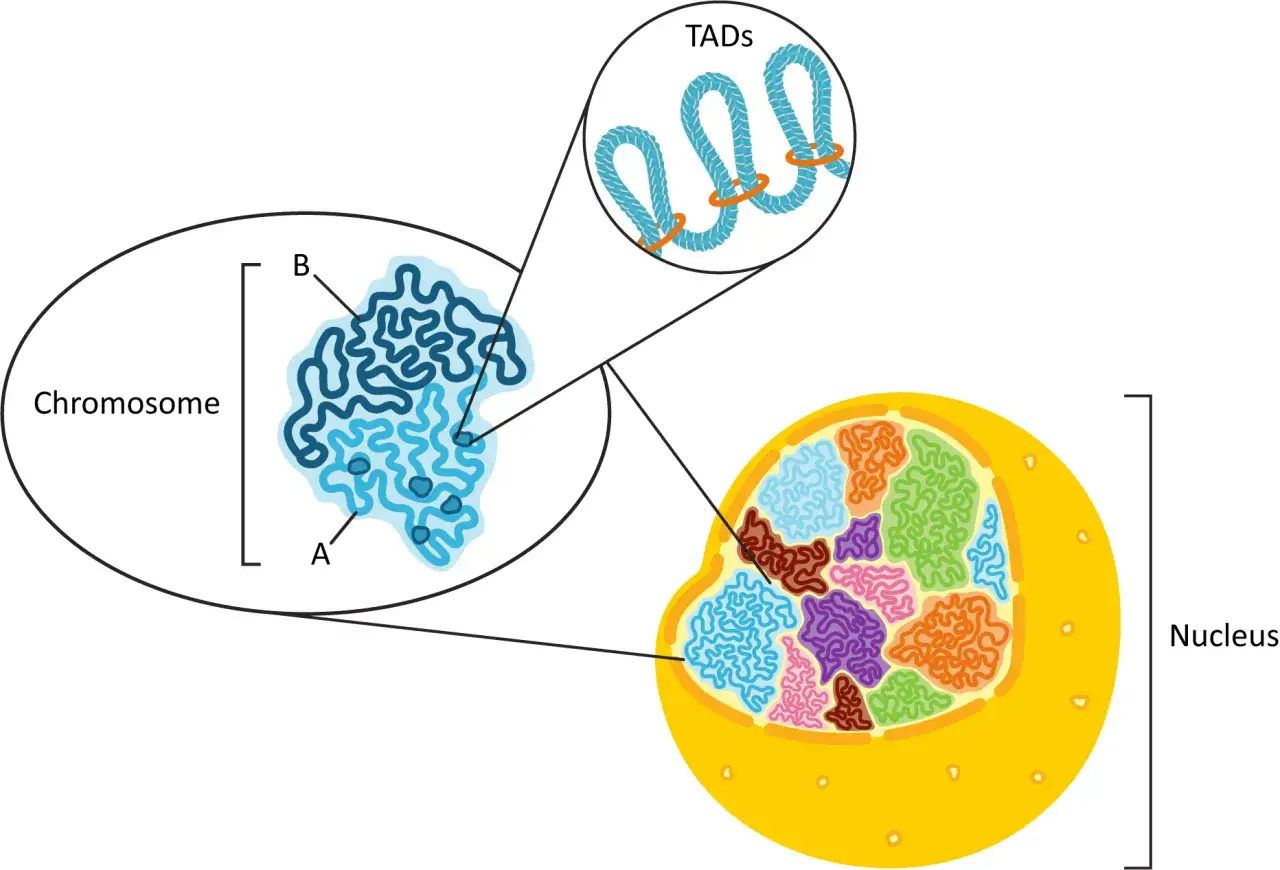
Jeśli spojrzeć tylko na liniową sekwencję, można przeoczyć najciekawszą część. Chromatyna układa się w pętle, domeny kontaktów i większe kompartmenty, które pomagają rozdzielić regiony aktywne od mniej aktywnych. To nie jest sztywna makieta, lecz ruchomy system, który zmienia się wraz ze stanem komórki.
W tym układzie ważną rolę odgrywają kompleksy SMC, czyli białkowe maszyny utrzymujące strukturę chromosomów. Najczęściej wymienia się cohesin i condensin. Cohesin pomaga tworzyć i stabilizować pętle DNA, a condensin jest szczególnie istotny przy silnym zagęszczaniu chromosomów w czasie podziału komórki. CTCF, białko wiążące DNA, często działa jak lokalny „znacznik granicy”, który porządkuje miejsca kontaktu między fragmentami chromatyny.
- Pętle chromatynowe zbliżają do siebie odległe fragmenty DNA, na przykład gen i jego element regulatorowy.
- Domeny TAD to obszary, w których DNA częściej kontaktuje się wewnątrz niż z sąsiednimi regionami.
- Kompartmenty A i B pomagają rozdzielać obszary bardziej aktywne od tych bardziej wyciszonych.
- Lamina jądrowa kotwiczy część chromatyny przy obwodzie jądra i wpływa na jej dostępność.
Warto tu zapamiętać jedną rzecz: położenie genu w jądrze nie jest wyrokiem, ale silną wskazówką. Regiony aktywne często znajdują się w bardziej otwartym środowisku, a regiony wyciszone bywają bliżej obwodu jądra lub w większych blokach heterochromatyny. Zdarzają się jednak wyjątki, więc nie traktuję tego jako sztywnej reguły podręcznikowej. Ta architektura zmienia się jednak w czasie, dlatego trzeba zobaczyć, co dzieje się podczas pracy samej komórki.
Co dzieje się z chromatyną podczas pracy komórki
Najbardziej praktyczne pytanie brzmi nie „jak to wygląda”, tylko „jak to działa w ruchu”. Odpowiedź jest prosta: komórka stale przebudowuje chromatynę. Korzysta przy tym z enzymów remodelujących, które zużywają ATP, oraz z białek opiekuńczych histonów, które pomagają zdejmować i ponownie układać nukleosomy.
Podczas transkrypcji
Gdy gen ma zostać aktywowany, lokalny fragment chromatyny musi się otworzyć. Pomagają w tym modyfikacje histonów, czynniki transkrypcyjne i kompleksy remodelujące. Dzięki temu RNA polimeraza może dotrzeć do matrycy DNA. To nie jest jednorazowe „odbezpieczenie”, tylko dynamiczna równowaga między otwarciem i ponownym uporządkowaniem regionu.
Podczas replikacji
W czasie kopiowania DNA nukleosomy muszą zostać częściowo rozebrane i złożone na nowo. To ważne, bo bez tego polimeraza replikacyjna nie przeszłaby przez gęsto upakowany materiał. Jednocześnie komórka stara się zachować informację epigenetyczną, czyli wzorzec modyfikacji histonów, który pomaga komórkom potomnym „pamiętać”, jaki mają typ i jak pracować.
Przeczytaj również: Od kiedy biologia w szkole podstawowej? Zmiany w programie nauczania
Podczas naprawy DNA
Uszkodzenie DNA wymaga szybkiego dostępu do danego miejsca. Wtedy chromatyna lokalnie się rozluźnia, a białka naprawcze zyskują przestrzeń do działania. To pokazuje, że zbyt ciasne upakowanie byłoby problemem, ale zbyt luźne też nie jest idealne. Komórka musi utrzymać precyzyjny balans między ochroną genomu a dostępnością jego fragmentów.
Właśnie dlatego upakowanie DNA nie jest biernym magazynowaniem, lecz ciągłym zarządzaniem ruchem, dostępem i bezpieczeństwem. Gdy ten porządek się psuje, skutki nie ograniczają się do jednego genu.
Co się psuje, gdy organizacja DNA zawodzi
Zaburzenia organizacji chromatyny widać na kilku poziomach. Najpierw pojawiają się błędy w ekspresji genów, później problemy z replikacją i naprawą, a w dalszej perspektywie niestabilność genomu. To właśnie dlatego defekty w białkach histonowych, remodelerach chromatyny czy kompleksach cohesinowych mają znaczenie kliniczne.
- Nieprawidłowa ekspresja genów może zmieniać różnicowanie komórek i ich funkcje.
- Gorsza naprawa DNA zwiększa ryzyko nagromadzenia mutacji.
- Niższa stabilność genomu sprzyja powstawaniu rearanżacji chromosomowych.
- Błędy rozwojowe mogą wynikać z zaburzonej regulacji całych programów genowych.
W praktyce bardzo często nie chodzi o jedną „złą” sekwencję, tylko o rozjechanie się całej logiki dostępu do DNA. Dlatego zaburzenia chromatyny są łączone z nowotworami, niektórymi zespołami rozwojowymi i ogólnie z problemami utrzymania tożsamości komórkowej. To mocny dowód, że organizacja DNA nie jest dodatkiem do biologii komórki, lecz jednym z jej fundamentów. Skoro tak, warto jeszcze odsiać kilka szkolnych skrótów myślowych, które często bardziej przeszkadzają niż pomagają.
Które szkolne skróty myślowe o chromatynie najbardziej mylą
Jeśli miałbym wskazać kilka rzeczy, które najczęściej upraszczają temat za mocno, wymieniłbym cztery. One naprawdę pomagają uporządkować wiedzę, szczególnie na początku, ale tylko wtedy, gdy nie traktuje się ich jak pełnego opisu rzeczywistości.
- „Chromatyna jest po prostu opakowaniem DNA” - to tylko połowa prawdy, bo chromatyna reguluje też dostęp do genów.
- „Heterochromatyna jest zawsze nieaktywna” - w praktyce jej rola bywa bardziej złożona, zwłaszcza w stabilizacji genomu.
- „30-nanometrowe włókno wyjaśnia wszystko” - to model pomocniczy, ale nie uniwersalny opis życia komórki.
- „Chromosom to zawsze ta sama struktura” - w interfazie i w mitozie chromosomy wyglądają i zachowują się zupełnie inaczej.
Najlepiej zapamiętać to tak: DNA nie jest po prostu zwinięte, tylko aktywnie zarządzane. Jeśli patrzy się na chromatynę przez ten pryzmat, łatwiej zrozumieć zarówno ekspresję genów, jak i podział komórki, a cały temat przestaje być zbiorem luźnych definicji. To właśnie ten sposób myślenia uważam za najuczciwszy i najbardziej użyteczny w biologii komórki.